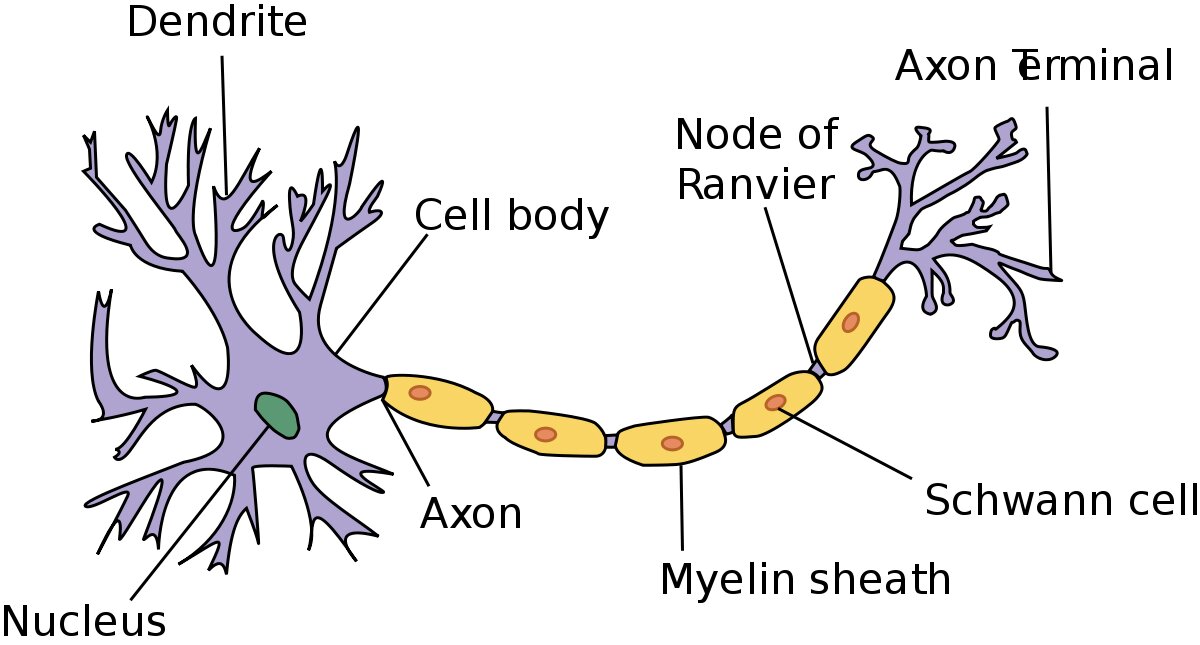
 Студентов-физиологов неизменно учат, что основная функция миелина — изоляция нервов. В частности, сделать потенциалы действия более эффективными, увеличив толщину мембраны и тем самым уменьшив ее электрическую емкость. Но эта грубая идея — эта аналогия на самом деле — не может быть правильной. Несмотря на протесты догматических нейробиологов, нейроны не являются электрическими устройствами, по крайней мере, не в том смысле, что электроны текут по проводам.
Студентов-физиологов неизменно учат, что основная функция миелина — изоляция нервов. В частности, сделать потенциалы действия более эффективными, увеличив толщину мембраны и тем самым уменьшив ее электрическую емкость. Но эта грубая идея — эта аналогия на самом деле — не может быть правильной. Несмотря на протесты догматических нейробиологов, нейроны не являются электрическими устройствами, по крайней мере, не в том смысле, что электроны текут по проводам.
Конечно, кажется, что между белками внутренней мембраны митохондрий глубоко внутри нейронов течет много электронных потоков (некоторые говорят, что до 50 ампер для всего тела), но эти токи не имеют ничего общего с распространением потенциала действия. Спайки представляют собой многогранные биофизические нарушения в аксонах. Они явно имеют ионный компонент в виде различных потоков натрия, калия, хлорида, кальция и, возможно, даже протонов, через каналы и насосы; однако электроны не являются здесь носителями тока или проводимости.
Так что же на самом деле делает миелин для аксона? Один популярный ответ заключался в том, что он обеспечивает некоторую энергетическую или трофическую поддержку, возможно, очень похожую на своего рода митохондрию, которая может производить АТФ посредством окислительного фосфорилирования через эктопически выраженные дыхательные комплексы. Существует увлекательная литература, описывающая присутствие предположительно функционирующего дыхательного комплекса V, F 1 F O -АТФ-синтазы, вне митохондрий в таких местах, как наружные сегменты палочек в сетчатке и в миелинизирующих клетках. Это может быть не так уж удивительно в свете присутствия нескольких видов АТФ-аз, обнаруженных в разных клеточных компартментах.
По-видимому, клеткам совершенно невозможно собрать полноценные дыхательные комплексы вне митохондрий из-за необходимости внутреннего конструирования и последующей вставки в мембрану чрезвычайно гидрофобных митохондриально экспрессируемых белков, а также интенсивного внутримитохондриального процессинга и созревания митохондриальных белков. белковые субъединицы ядерного происхождения (см., например, эту критику аллотопической экспрессии митохондриальных белков). Тем не менее, есть и другие способы, которыми эти белки могут попасть на плазматическую мембрану после того, как они были собраны. Творческое разрушение частей митохондрий с образованием всевозможных одно- и двухслойных почкующихся везикул, вероятно, могло бы переправлять комплексы на границу и дальше.
В текущем выпуске Open Biology Королевского общества Alessandro Morelli et al. представили интригующие доказательства того, что миелиновые оболочки с концентрическими мультиламеллярными структурами обладают сходной биоэнергетикой с цианобактериальными тилакоидами. В дополнение к множеству общих эклектичных молекулярных усовершенствований обе структуры также разделяют очевидную функцию подачи питательных веществ, потенциально включая АТФ, полученный из АТФ-синтазы, в центральное сердце сложной многослойной структуры. Никто не утверждает, что сам миелин был получен из мембран тилакоидов, поскольку это казалось бы таксонометрически невозможным, но эти наблюдения могут представлять собой поучительный пример конвергентной эволюции для выполнения какой-то фундаментально похожей задачи.
Помимо грубой структуры и образования АТФ, есть и другие ключи к разгадке этой общей базовой функции. Например, плотно упакованные концентрические липиды надежно выглядят как оптимальная природная конструкция для растворения и секвестрации наибольшего количества газа в определенном объеме. Липиды, особенно нейтральные липиды, могут удерживать примерно в пять раз больше газа, чем вода. В этом случае цианобактерии будут наиболее заинтересованы в растворении CO 2 для построения углеводов и азота для фиксации, в то время как миелин, несомненно, будет искать O 2 . Ткани мозга не могут позволить себе роскошь (как и другие ткани с высоким уровнем дыхания, такие как мышцы) иметь миоглобин с высоким сродством для захвата O 2 из циркулирующего гемоглобина.
Может быть интересно сравнить коронарные срезы мозга китообразных с человеческим в свете значительно более тонкой и извитой коры китообразных, которая развилась под селективным давлением условий, лишенных кислорода. Кора китообразных, вероятно, может содержать большое преобладание крупных проекционных нейронов глубокого слоя по сравнению с их тонкими верхними слоями, чтобы максимизировать количество аксонов, доступных для производства белого вещества. Беглый взгляд на мозг дельфинов на изображениях Google не подтверждает это так или иначе, однако более исчерпывающий учет может пролить свет. Точно так же спермацет кашалота, который предположительно формирует лучи и фокусирует входящие или исходящие сигналы эхолокации, может подрабатывать в качестве кислородного питания. Их воск по преимуществу, т.е.цетилпальмитат представляет собой жирную кислоту C16, которая этерифицирует спирт и может иметь значение для поглощения кислорода. Любопытно, что концентрические мультиламеллярные структуры также образуются в легочном сурфактанте, ключевом элементе хорошего O.2 абсорбция легочными альвеолами. Внешний слой легочных эпителиальных клеток также имеет сходные образования пластинчатого тела.
Чтобы преодолеть отрицательно заряженные силы отталкивания фосфолипидов в мультиламеллярных структурах, природа, по-видимому, единогласно пошла на галактолипиды. Эти липиды, содержащие галактозу в качестве сахарной группы, являются фаворитами среди липидов растительных мембран, где они заменяют фосфолипиды, по-видимому, для сохранения фосфата для других важных процессов. Галактолипиды составляют около 70 % липидов тилакоидов цианобактерий, до 80 % тилакоидов растений и около 30 % миелина. Также было показано, что миелиновые галактолипиды необходимы для правильного формирования перехватов Ранвье.
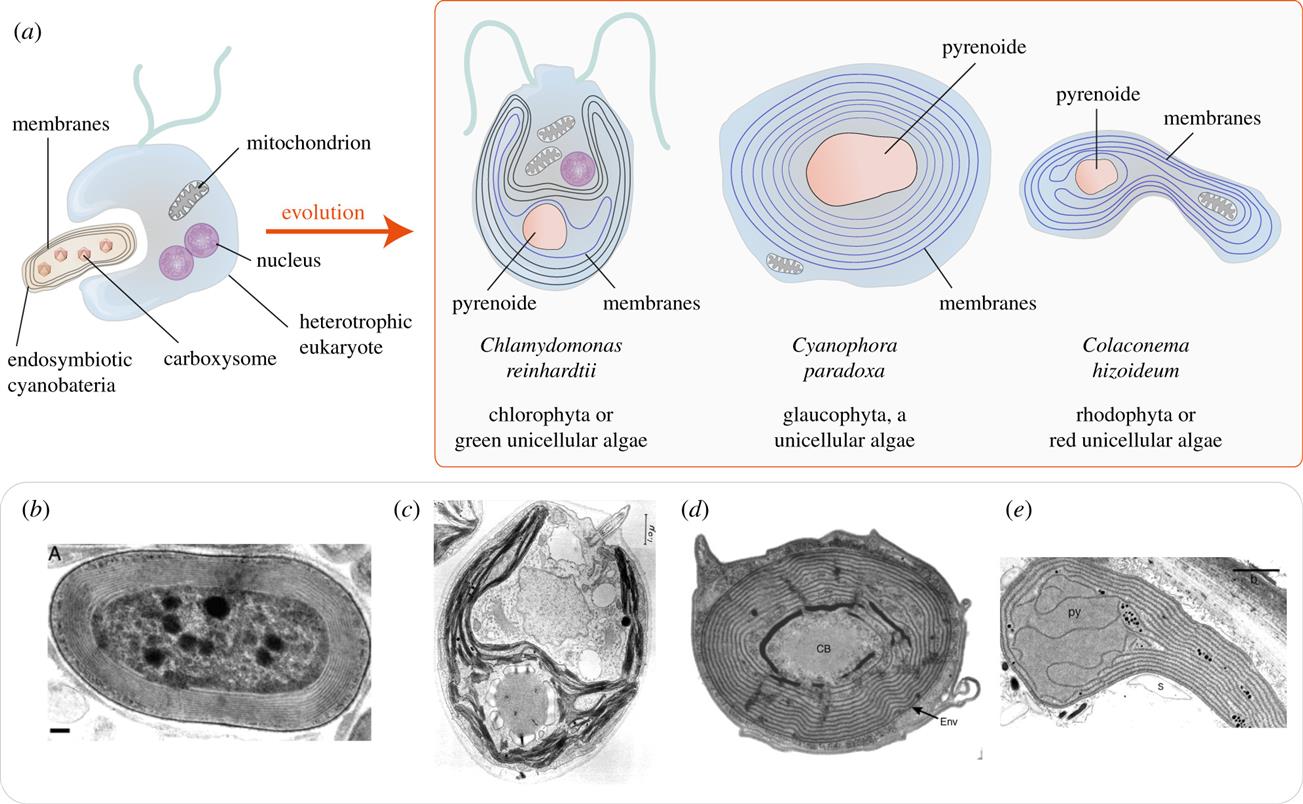
Другое сходство с тилакоидами — близкая гомология белковых последовательностей неселективных ионных каналов в тилакоидах с вездесущими потенциалзависимыми анионными каналами (VDAC), обильно экспрессируемыми в митохондриях и других мембранных структурах. В миелине VDAC потенциально может тетрамеризоваться в комплексы, которые образуют центральную пору диаметром примерно 1,3 нм, примерно того же размера, что и пора ионного канала тилакоидов. У зеленой одноклеточной водоросли Chlamydomonas reinhardtii эти каналы соединяют стопки тилакоидов с важной структурой, называемой пиреноидом. Бикарбонат, НАДФН и АТФ проходят через поры к центральному пиреноиду, чтобы питать цикл Кальвина-Бенсона.
Для включения СО 2 в органические соединения требуется очень высокая концентрация фермента RuBisCo . Будучи самым распространенным белком во всей нашей биосфере, RuBisCo демонстрирует лишь скромную каталитическую эффективность. Это, вероятно, причина того, что плотные ядра комплексов RuBusCo, образующих карбоксисомы, находятся глубоко внутри центров концентрических ламеллярных структур. Для транспортировки CO 2 к рабочему концу тилакоидов требуется карбоангидраза для образования промежуточного бикарбоната, как это происходит у нас. Карбоангидраза необходима в митохондриях для превращения CO 2 , полученного в результате цикла Кребса, в бикарбонат. Еще одно место, где карбоангидраза находится миелиновая оболочка, что указывает на метаболическую роль в обращении с этими газообразными веществами.
Митохондрии содержат обширные мембранные компартменты, известные как кристы. В норме эти элементы образуют параллельные стопки, спорадически связанные с наружной мембраной соединениями трубчатых крист. При определенных обстоятельствах эти кристы могут полностью перестраиваться в концентрические луковичные мембраны. Многие манипуляции, как естественные, так и искусственные, заставляют митохондрии принимать концентрические кристы, включая изменения в экспрессии АТФ-синтазы. Некоторое время назад несколько исследователей предположили, что миелин действует как митохондрия. Теоретическое возражение против этой концепции позже утверждало, что если ошибочная АТФ-аза найдет свое место в миелине, то в лучшем случае она сможет действовать только в обратном направлении, расщепляя АТФ.
Эти авторские расчеты протонной движущей силы через миелиновую мембрану были основаны на известных значениях pH и мембранного потенциала олигодендроцита, а также на нескольких предположениях об основной конфигурации и полярности АТФ-синтазы в мембране. Теоретические доказательства того или иного метаболизма, например количества энергии, необходимой нейронам, общеизвестно сложны — природа неизменно удивляет тем, как много она может сделать с таким небольшим количеством энергии. Например, те же самые исследователи, бросающие тень на миелиновую гипотезу, также предприняли благородные попытки рассчитать потребность в энергии для спайковых аксонов. В то время как многие источники скрытой энергии также доступны для аксона, например, генераторы АТФ, прикрепленные к пузырькам в форме GADPH аксоны делают гораздо больше, чем спайки. На самом деле всплеск может быть относительно простым по сравнению с более физическими процессами, такими как транспорт и рост.
Питер Митчелл, лауреат Нобелевской премии, ввел понятие H+-движущей силы (обычно около 250 мВ), которая будет состоять из трансмембранной разности электрических потенциалов (Δψ) плюс разность pH между двумя водными фазами (ΔpH). Однако в разговоре с Алессандро, ныне живущим пионером, ощутимо связанным с той благодатной эпохой и все еще весьма активным сегодня, быстро становится очевидным, что не все может быть хорошо в хемиосмотической стране. Было замечено, что поверхность биологической мембраны отделена от объемной водной фазы упорядоченными молекулами воды, образующими электростатический барьер, который для Н+ составляет около 120 мэВ. Таким образом, любой расчет значения pmf на разных поверхностях далеко не прост, если принять во внимание реальную биологию.
Природу свободных протонов в мембранах и вокруг них также трудно интуитивно понять. Особенно высокая концентрация протонов, вероятно, не очень полезна для мембран. Благодаря более совершенным методам физические переменные в таких структурах, как митохондрии, теперь можно измерять с большей пространственной и временной точностью, часто с поразительными результатами. Глубоко внутри матрикса температура буквально пылает, кальций выжигает искры, а потенциалы митохондриальной мембраны меняются в пределах порядка скорости спайков самого родительского аксона.
По материалам зарубежной прессы